Характеристика половых хромосом. Хромосомы
У позвоночных животных очень часто ключевую роль в определении пола играют половые хромосомы. Если у низших позвоночных в определении пола также нередко участвуют факторы окружающей среды, то у птиц и млекопитающих определение пола строго хромосомное. Как правило, в кариотипе есть две половые хромосомы: X и Y у млекопитающих (самки имеют кариотип XX, самцы - XY) или Z и W у птиц (ZW у самок и ZZ у самцов). Впрочем, иногда половых хромосом в кариотипе больше двух. Абсолютным рекордсменом по этому показателю долгое время считался утконос: из 52 его хромосом в качестве половых функционируют 10. Однако недавно невзрачная южноамериканская лягушка, известная как пятипалый свистун (Leptodactylus pentadactylus ), уверенно утерла ему нос: из 22 ее хромосом более половины (а именно, 12) являются половыми! Наша статья посвящена этому любопытнейшему открытию.
У многих низших позвоночных - рыб, амфибий и рептилий - как таковых половых хромосом, морфологически отличающихся от остальных хромосом (аутосом), нет. При этом у млекопитающих и птиц обязательно есть половая хромосома , утратившая значительную часть генов - Y-хромосома в случае млекопитающих и W-хромосома в случае птиц. В тех случаях, когда половые хромосомы все-таки есть, они обычно представлены одной парой: XX♀:XY♂ или ZZ♂:ZW♀. Причины, по которым у низших позвоночных нет морфологически выделяющихся (гетероморфных) половых хромосом, не совсем ясны. Существует два предположения на этот счет. Согласно одному из них, мутации в генах, участвующих в определении пола, происходят так часто, что хромосомы просто не имеют возможности начать терять их из-за необходимости все время устранять мутации в этих чрезвычайно важных генах, возвращаясь к исходному состоянию. Вторая гипотеза предполагает, что дегенерации половых хромосом препятствуют многочисленные акты рекомбинации , в ходе которых утерянные фрагменты восстанавливаются.
Однако в биологии нет правил без исключений. Известны примеры амфибий, имеющих несколько гетероморфных половых хромосом. Например, у лягушек Strabomantis biporcatus и Pristimantis riveroi определение пола происходит по схеме X 1 X 1 X 2 X 2 ♀:X 1 X 2 Y♂. В 2016 году в Швеции была найдена популяция травяных лягушек (Rana temporaria ), у которых имеется две Х-хромосомы и две Y-хромосомы. Большинство примеров наличия нескольких половых хромосом относится к млекопитающим. Например, у утконоса имеется 10 половых хромосом, из которых 5 являются Х-хромосомами и 5 - Y-хромосомами .
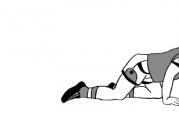
Рисунок 1. Кольцевая структура, формирующаяся при мейозе у самцов пятипалого свистуна. Отчетливо видны 12 хромосом, образующих кольцо. ДНК окрашена синим , красным выделены теломеры.
О том, что такое флуоресцентная микроскопия и как она работает, можно прочитать в статье «12 методов в картинках: микроскопия » .

Рисунок 2. Кольцевые структуры в сперматоцитах двух самцов пятипалого свистуна. Хромосомы окрашены по Гимзе. Масштабная линейка 5 мкм.
Кольцевая структура в сперматоцитах пятипалого свистуна состоит из 12 хромосом, при этом полный кариотип этой лягушки включает 22 хромосомы. Таким образом, пятипалый свистун - единственный известный на данный момент вид позвоночных, в кариотипе которого половых хромосом больше, чем аутосом. Ученые предполагают, что Y-хромосома пятипалого свистуна претерпела целых семь
1. Какие хромосомы называются половыми?
Ответ. Половые хромосомы - это пара хромосом, которые различаются у мужских и женских особей одного биологического вида. У одного из полов это, как правило, две одинаковые крупные хромосомы (Х-хромосомы, генотип XX); у другого - одна Х-хромосома и одна меньшая по размеру Y-хромосома (генотип XY). У некоторых видов мужской пол формируется при отсутствии одной половой хромосомы (генотип Х0).
2. Какие организмы называются гермафродитами?
Ответ. Гермафродит – организм, у которого имеются мужские и женские гонады, образующие половые клетки у одной особи. Такой гермафродитизм встречается у плоских и кольчатых червей. Это –истинный гермафродитизм. Разновидностью его может быть гермафродитизм моллюсков, половая железа которых, в зависимости от возраста и условий существования, периодически продуцирует то мужские, то женские гаметы. В случае ложного гермафродитизма у одной особи развиваются наружные половые органы и вторичные признаки обоих полов, а гонады – одного пола (мужские или женские).
3. Какие болезни называются наследственными?
Ответ. Наследственные болезни - это болезни, обусловленные изменением генотипа (т.е. мутациями). Они не всегда передаются от родителей к детям. Многие наследственные заболевания не могут наследоваться (передаваться из поколения в поколение), так как они снижают жизнеспособность больного или вызывают бесплодие. Могут возникать у детей здоровых родителей в результате новой мутации. Например, у здоровых, как правило, родителей рождается ребенок с синдромом Дауна. С другой стороны, некоторые эндемичные заболевания наблюдаются у родителей и детей. Создается впечатление наследования, но заболевания не являются наследственными (например, эндемичный зоб).
Вопросы после §45
1. Какие типы хромосом вам известны?
Ответ. Хромосомы подразделяют на половые и неполовые (аутосомы). Половые хромосомы - это пара хромосом, которые различаются у мужских и женских особей одного биологического вида. У одного из полов это, как правило, две одинковые крупные хромосомы (Х-хромосомы, генотип XX); у другого - одна Х-хромосома и одна меньшая по размеру Y-хромосома (генотип XY). У некоторых видов мужской пол формируется при отсутствии одной половой хромосомы (генотип Х0). Аутосомы - это пары хромосом, которые идентичны у особей одного биологического вида, относящихся к разным полам. Число пар аутосом равно числу пар хромосом в генотипе минус единица (одна пара половых хромосом). Так, у человека 22 пары аутосом, у дрозофилы - 3 пары. Всем аутосомам каждого биологического вида даны порядковые номера в соответствии с их размером (первая - самая большая; последняя - самая короткая и, следовательно, несущая меньше всего генов)
2. Что такое гомогаметный и гетерогаметный пол?
Ответ. Гомогаметным называют пол, который формирует гаметы одного типа по половым хромосомам (генотип XX). Гетерогаметный пол в процессе гаметогенеза образует гаметы двух типов по половым хромосомам (генотип XY либо Х0). У человека гомогаметен женский пол, гетерогаметен мужской (генотип XY)
3. Как наследуется пол у млекопитающих?
Ответ. У мужских и женских организмов все пары хромосом, кроме одной, одинаковы и называются аутосомами, а одна пара хромосом, называемых половыми, – у самцов и самок различается. У самцов и самок половые хромосомы различаются: у самок – по две Х-хромосомы, а у самцов X и Y. Пол будущей особи определяется во время оплодотворения. Если сперматозоид содержит Х-хромосому, то из оплодотворённой яйцеклетки разовьётся самка (XX), а если в сперматозоиде содержалась половая Y-хромосома – то самец (XY).
4. Какие другие варианты хромосомного и нехромосомного определения пола у живых организмов вам известны? Приведите конкретные примеры.
Ответ. У птиц и рептилий – гомогаметны самцы (XX), а самки – гетерогаметны (XY). У некоторых насекомых у самцов в хромосомном наборе лишь одна половая хромосома (Х0), а самки – гомогаметны (XX).
У пчёл и муравьёв половых хромосом нет, и самки имеют в клетках тела диплоидный набор хромосом, а самцы, развивающиеся партеногенетически (из неоплодотворённых яйцеклеток), – гаплоидный набор хромосом. Естественно, что в этом случае развитие сперматозоидов у самцов идёт без мейоза, так как уменьшить число хромосом менее гаплоидного набора невозможно.
У крокодилов половые хромосомы не обнаружены. Пол зародыша, развивающегося в яйце, зависит от температуры окружающей среды: при высоких температурах развивается больше самок, а в том случае, если прохладно, – больше самцов.
5. Мужской или женский пол у человека является гетерогаметный?
Ответ. У человека гетерогаметный мужской организм (XY).
6. Имеются ли различия по числу хромосом между маткой и рабочими особями медоносной пчелы?
Ответ. Каждая клетка медоносной пчелы (матки и рабочие особи) имеется в ядрах клеток по 32 хромосомы.
Однако у медоносных пчел имеются существенные отличия от общепринятого механизма передачи наследственности родителей своим потомкам и определение пола особей. Если у всех сельскохозяйственных животных пол особи определяется определенными половыми хромосомами, то у медоносных пчел пол особи определяется по-другому: при оплодотворении яиц из них образуются самки (матки и рабочие пчелы), а из неоплодотворенной яйцеклетки матки развиваются самцы (трутни).
Таким образом, женские особи семьи являются диплоидными организмами (имеют 32 хромосомы), а мужские особи (трутни) - гаплоидными (имеют в своих клетках по 16 хромосом).
Половые хромосомы, в отличие от аутосом, обозначаются не порядковыми номерами, а буквами X, Y, W или Z, причём отсутствие хромосомы обозначается цифрой 0. При этом один из полов определяется наличием пары одинаковых половых хромосом (гомогаметный пол, XX или WW), а другой - комбинацией двух непарных хромосом или наличием только одной половой хромосомы (гетерогаметный пол, XY, WZ или X0). У человека, как и у большинства млекопитающих, гомогаметный пол - женский (XX), гетерогаметный пол - мужской (XY). У птиц, напротив, гетерогаметный пол - женский (WZ), а гомогаметный - мужской (WW). У амфибий и рептилий имеются виды (например, все виды змей) с гомогаметными самцами и гетерогаметными самками, а некоторые черепахи (крестогрудая черепаха Staurotypus salvinii и черная пресноводная черепаха Siebenrockiella crassicollis), наоборот, имеют гетерогаметных самцов и гомогаметных самок. В некоторых случаях (у утконоса) пол определяется не одной, а пятью парами половых хромосом
Рисунок 13. Карта Х-хромосомы человека
На стрекозах показано, что форма XY эволюционно более поздняя, чем ХО. Другая точка зрения - половые хромосомы произошли от обычной пары аутосом, несущей гены, определяющие пол. Поэтому у одних видов (более примитивных) Y-хромосома такая же по размерам, как и Х-хромосома, конъюгирует с ней полностью или частично, участвует в кроссинговере. А у других видов - она маленькая, с Х-хромосомой соединяется конец в конец, без кроссинговера. В процессе эволюции Y- хромосома почему-то теряет активные гены, деградирует и исчезает, потому форма XY предшествует ХО.

Рисунок 14. Половые хромосомы (Х и Y)
Y-хромосома - самая вариабельная хромосома генома. У человека она генетически почти пустая (ген волосатости ушей и перепонок между пальцами ног). У других видов может содержать много активных генов - у гуппи - около 30 Y-генов окраски самцов (и только 1 аутосомный ген).
Y-хромосома Drosophila. Содержит 9 генов: 6 определяют фертильность самцов, 3 bobbed кластер генов рРНК. Активность bb генов приводит к формированию ядрышка. Ядрышкообразующий bb ген есть и в Х-хромосоме - сайт спаривания Х и Y хромосом - сайт collohaes. Ответственными за конъюгацию являются короткие последовательности нуклеотидов (240 п.н.), расположенные между генами рРНК в Х и Y - хромосомах. Удаление bb локуса - нет конъюгации половых хромосом. Ещё один ген - crystal - влияет на поведение хромосом в мейозе. Его делеция - нарушается расщепление хромосом в мейозе.
У дрозофилы 6 факторов фертильности самцов. Из них 3 очень большие - занимают по 10% Y- хромосомы каждый, т.е. по 4000 т.п.н.
В составе ДНК Y-хромосомы 2 типа последовательностей:
Y - специфичные - семейства из 200-2000 копий, организованы в кластеры тандемно повторенных единиц длиной 200-400 п.н. Расположены, вероятно, в петлях.
Y-ассоциированные (встречаются в других хромосомах).
Y-хромосома человека
Y-хромосома является наименьшей по размеру из 24 хромосом у человека и содержит около 2-3% ДНК гаплоидного генома, составляя приблизительно 51 Mb. Из всего объема ДНК Y-хромосомы на данный момент секвенировано 21.8 Mb. Короткое плечо Y-хромосомы (Yp) содержит примерно 11 Mb, а длинное плечо (Yq) - 40 Mb ДНК, из которых около 7 Mb приходятся на эухроматиновую часть Yq и около 3 Mb ДНК на центромерную область хромосомы. Большая часть (~60%) длинного плеча Y-хромосомы представляет собой функционально неактивный гетерохроматин, имеющий размер около 24 Mb. В Y-хромосоме выделяют несколько областей: псевдоаутосомные области (PARs); - эухроматиновую область короткого плеча (Yp11); - эухроматиновую область проксимальной части длинного плеча (Yq11); - гетерохроматиновую область дистальной части длинного плеча (Yq12); - область прицентромерного гетерохроматина.
Y-хромосома содержит около 100 функциональных генов. Из-за наличия на Х и Y-хромосомах (на теломерах) гомологичных PAR-регионов, половые хромосомы регулярно конъюгируют и рекомбинируют участками этих регионов в зиготене и пахитене профазы I мейоза. Однако большая часть (~95%) Y-хромосомы не принимает участия в рекомбинации, и поэтому называется нерекомбинирующей областью Y-хромосомы (NRY - Non Recombinant Region Y chromosome).
Гетерохроматиновая область длинного плеча Y-хромосомы является генетически инертной и содержит различные типы повторов, в том числе высокоповторяющиеся последовательности двух семейств DYZ1 и DYZ2, каждый из которых представлен приблизительно 5000 и 2000 копиями соответственно.
На основе сравнительного анализа генов гоносом X и Y в Y-хромосоме выделяют три группы генов:
1. PAR-гены (PAR - Pseudoautosomal Region; гены псевдоаутосомных областей PAR1 и PAR2), локализованные в теломерных областях Y-хромосомы;
2. X-Y гомологичные гены, локализованные в нерекомбинирующих областях Yp и Yq;
3. 3. Y-специфичные гены, расположенные в нерекомбинирующих областях Yp и Yq.

Рисунок 15. Y-хромосома
Первая группа представлена генами псевдоаутосомных областей (регионов). Они являются идентичными для X- и Y-хромосом и наследуются как аутосомные гены. PAR1-регион расположен на конце короткого плеча Y-хромосомы, он больше по размеру, чем PAR2-регион, локализованный на конце длинного плеча Y-хромосомы, и его размер приблизительно оценивается в 2,6 Mb. Так как делеции PAR1 приводят к нарушениям конъюгации гоносом во время мейоза у мужчин и могут привести к мужскому бесплодию, предполагается, что PAR-регионы имеют существенное значение для нормального протекания сперматогенеза у мужчин.
Вторая группа генов содержит X-Y-гомологичные, но не идентичные гены, которые локализованы в нерекомбинирующих районах Y-хромосомы (на Yp и Yq). В нее включены 10 генов, представленных на Y-хромосоме одной копией, большинство из них экспрессируются у человека во многих тканях и органах, включая яички и предстательную железу. До сих пор неизвестно, являются ли эти X-Y-гомологичные гены функционально взаимозаменяемыми.
Третью группу генов составляют 11 генов, которые расположены в нерекомбинирующем районе Y-гоносомы (NRY). Все эти гены, за исключением гена SRY (Sex-Determining Region Y Chromosome, пол-детерминирующий регион Y-хромосомы), представленного одной копией, являются мультикопийными, и их копии расположены на обоих плечах Y-хромосомы. Некоторые из них являются генами-кандидатами на AZF-фактор (Azoospermia factor, или фактор азооспермии).
О точных функциях большинства этих генов известно мало. Продукты, кодируемые генами нерекомбинирующего региона Y-хромосомы, выполняют различные функции, например, среди них имеются факторы транскрипции, цитокиновые рецепторы, протеинкиназы и фосфатазы, которые могут влиять на клеточную пролиферацию и/или передачу сигналов в клетке.
На длинном плече Y-хромосомы расположен AZF (Azoospermia Factor) локус - содержит гены, контролирующие процесс дифференцировки половых клеток, т.е. сперматогенез. В данном локусе выделяют 3 региона - a (800 т.п.н.), b (3,2 млн пн), c (3,5 млн. пн). Микроделеции участков данного локуса являются одной из основных генетических причин мужского бесплодия. Микроделеции длинного плеча Y-хромосомы обнаруживаются у 11% мужчин с азооспермией и у 8% мужчин с олигозооспермией тяжелой степени. При делеции всего с-региона AZF локуса возможно возникновение блока в митозе и мейозе при сперматогенезе; на гистологических препаратах у таких больных в большинстве семенных канальцев отсутствуют половые клетки.
Для Y-хромосомы характерны специфические черты, резко отличающие ее от других хромосом человека: 1) обедненность генами;
2) обогащенность повторяющимися блоками нуклеотидов. Присутствие значительных гетерохроматиновых районов;
3) наличие области гомологии с Х-хромосомой - псевдоаутосомальной области (PAR) (Черных, Курило, 2001).
Y-хромосома, как правило, не велика - 2-3% гаплоидного генома. Тем не менее, кодирующей способности ее ДНК у Homo sapiens достаточно по крайней мере для нескольких тысяч генов. Однако у этого объекта в Y-хромосоме выявляется всего около 40 обогащенных ГЦ-парами так называемых ЦрГ-островков, обычно фланкирующих большинство генов. Реальный же список генетических функций, связанных с этой хромосомой, вдвое меньше. Фенотипическое влияние этой хромосомы у мышей ограничено весом тестисов, уровнем тестостерона, серологического HY-антигена, чувствительностью органов к андрогенам и сексуальным поведением. Большая часть генов этой хромосомы имеет X-хромосомные аналоги. Большинство Y-хромосомных последовательностей гомологичны ДНК Х-хромосомы или аутосом и лишь часть из них строго уникальна.
Наличие псевдоаутосомальных областей, обеспечивающих мейотическое спаривание и рекомбинацию, обычно рассматривается как необходимое условие фертильности. Интересно, что размер участка мейотического спаривания существенно длиннее PAR. У человека имеются два псевдоаутосомальных района на вершине короткого и длинного плеч Х-хромосомы. Однако, только для первого из них установлены облигатный обмен в мейозе, наличие хиазм, влияние на фертильность.
Высказано предположение о происхождении половых хромосом млекопитающих от предковой аутосомы в результате независимых циклов: добавление - рекомбинация - деградация. PAR, по такой терминологии, представляет собой лишь как бы реликт такого последнего добавления. Далее происходят деградация и потеря соответствующих Y-хромосомных частей и инактивация Х-хромосомы. Все гены, представленные в Y-хромосоме, или имеют реальную селективную ценность (например, SRY), или находятся на пути исчезновения. Каждый Y-хромосомный ген, быстро дивергирующий, амплифицирующийся или склонный к исчезновению, имеет своего гомолога в Х-хромосоме, более консервативного и активного у обоих полов. Так, Sox3, предполагаемый X-хромосомный гомолог SRY, кодирует почти идентичные продукты у человека, мыши и сумчатых, экспрессируется в нервной системе обоих полов. SRY быстро дивергирует и активен только в гонадном бугорке. Этот Y-хромосомный ген подвергается амплификации у многих мышей и крыс.
Таким образом, Y-хромосома, единственная в геноме млекопитающих, не работает непосредственно на реализацию фенотипа. Ее генетическая значимость связана с преемственностью между поколениями, в частности с контролем гаметогенеза, первичной детерминацией пола. Жесткий отбор действует только на немногие ее гены, остальная ДНК более пластична.
Половое размножение свойственно всем живым организмам за исключением тех, которые вторично утратили половой процесс. Определение и развитие пола — сложный процесс, который детерминирован генетически, т.е. находится под контролем генов, а также подвержен влиянию внешней среды.

В животном мире господствует раздельнополость, т.е. существуют два типа ясно различающихся в половом отношении организмов — самцы и самки. Различия между ними очень глубокие и затрагивают не только органы, непосредственно участвующие в половом размножении. Половые различия сопровождаются заметными различиями в росте, обмене веществ, инстинктах, а также в тех признаках, которые подвержены воздействию половых желез, например, гребни, рога, волосы, оперение.
Гермафродитизм у животных в норме встречается только у немногих видов, например у червей.
У растений, наоборот, преобладает гермафродитность . Половые различия у растений выражены менее резко, чем у животных. Для растений характерны переходы от обоеполости к однополости, частые аномалии в развитии генеративных органов, изменение пола под влиянием внешних условий.
Определение пола у разных организмов может происходить на разных стадиях жизненного цикла.
Пол зиготы может предопределяться еще в процессе созревания женских гамет — яйцеклеток. Такое определение пола называется прогамным , т.е. оно происходит до оплодотворения. Прогамное определение пола обнаружено у коловраток и кольчатых червей. Яйцеклетки этих животных в результате неравномерного распределения цитоплазмы в процессе оогенеза различаются по размеру. Из крупных яйцеклеток после определения развиваются только самки, из мелких — только самцы.
Наиболее распространенным типом определения пола является сингамное , т.е. определение пола в момент слияния женских и мужских гамет. Оно встречается у млекопитающих, птиц, рыб и др.
Известен также третий тип определения пола — эпигамное , которое происходит на ранних стадиях индивидуального развития особи (например, у морского червя Bonelia viridis).
У большинства животных и раздельнополых растений основную роль в определении пола играют половые хромосомы . Еще в начале ХХ в. (1902 г., McClung) было установлено, что у некоторых насекомых (клоп Protenor) самцы образуют два типа сперматозоидов: один тип — с лишней хромосомой, второй — без нее. У самцов клопа Protenor в одних сперматозоидах было 7 хромосом, в других — 6. Непарную хромосому назвали половой хромосомой, в отличие от остальных — аутосом . В соматических клетках самца содержится 13 хромосом, одна из которых Х-хромосома (12A+X), в соматических клетках самки — 14 хромосом (12A+XX). Женский пол клопа является гомогаметным, так как образует гаметы одного типа (6A+X), а мужской — гетерогаметным и образует два типа гамет (6A+X) и (6А+0). Такой тип определения пола, при котором самки имеют кариотип ХХ , а самцы — Х0 , назван Protenor-типом. Он описан у большинства прямокрылых насекомых, жуков, пауков, многоножек и нематод.
Вслед за Protenor-типом был открыт другой тип определения пола, который характерен для млекопитающих, многих рыб, амфибий и ряда растений. Впервые он был описан у клопа Lygaeus turcicus и получил название Lygaeus-типа. При этом типе определения пола имеются два вида половых хромосом: Х и Y . Самки имеют две хромосомы, а самцы одну Х-хромосому и непарную ей Y-хромосому. Обозначение половых хромосом буквами X и Y отражает их форму, которую они имеют в профазе мейоза в результате отталкивания хроматид, соединенных только в области первичной перетяжки.
Женский пол при типе Lygaeus является гомогаметным, мужской — гетерогаметным.
У птиц, некоторых видов бабочек и рыб тип определения пола — обратный Lygaeus, т.е. гомогаметным является мужской пол. В этом случае для обозначения половых хромосом используют другие буквы: ♀ZW, ♂ZZ.
У моли описан тип — обратный Protenor, т.е. ♀Х0, ♂ХХ.
Особый тип определения пола характерен для пчел. Здесь разница между полами затрагивает не одну пару хромосом, а весь набор. Самки пчел — диплоидны, а самцы — гаплоидны, так как женские особи развиваются из оплодотворенных яйцеклеток, мужские особи — в результате партеногенеза.
Хромосомный механизм определения пола у растений был впервые определен у печеночного мха — Sphaerocarpus в ходе тетрадного анализа. Из четырех спор, образующихся в результате мейотического деления материнской клетки, две дают начало женским растениям, а две другие — мужским. Поскольку хромосомы мха Х и Y морфологически легко различимы, было установлено, что женские растения имеют кариотип 7А + Х, а мужские — 7А + Y. Диплоидный спорофит, который образуется в результате оплодотворения, имеет кариотип 14А + XY.
Гетероморфные пары хромосом обнаружены у мужских растений дремы, конопли, щавеля, хмеля и др. Определение пола у них соответствует типу Lygaeus. У земляники гетерогаметным (XY ) является женский пол, мужской — гомогаметным.

Половые хромосомы отличаются от аутосом поведением в профазе мейоза. Во время гаметогенеза они находятся в сильно спирализованном состоянии и редко объединяются в биваленты. Тем не менее они обладают сегментной гомологией и проявляют тенденцию к частичной коньюгации.
X и Y -хромосомы различаются по форме, величине и генному составу. Х-хромосома чаще всего относится к разряду крупных хромосом с большим генетическим объемом. У дрозофилы Х-хромосома — самая крупная в наборе. У человека Х-хромосома относится к разряду средних метацентриков, с нарушением ее структуры связан ряд тяжелых наследственных патологий (синдромов). Мужскую половую хромосому характеризует обедненность генами и, соответственно, низкая генетическая активность, а иногда и полная инертность. У человека с помощью молекулярно-генетических методов в Y-хромосоме выявлено около 40 генов. Однако реальных генетических функций еще меньше. В частности, в Y-хромосоме лежит мутация, отвечающая за малоприятный для мужчин признак — волосатость ушей. У дрозофилы Y-хромосома практически не оказывает никакого влияния на развитие пола.
У растений Y-хромосома также ведет себя по-разному: у одних она играет активную роль в определении пола, у других — является инертной. Например, Y-хромосома Milandrium alba (дрема) имеет сегменты, потеря которых ведет к нарушению нормального процесса развития пола и, как следствие, к мужской или женской стерильности. У Rumex acetosa Y-хромосома генетически инертна. У некоторых растений активность Y-хромосомы настолько высока, что особи YY оказываются жизнеспособными, как у аспарагуса, в то время как у других видов подобные особи не выживают.
Если гены, детерминирующие признаки, находятся в половых хромосомах, то их наследование не подчиняется законам Менделя. Распределение этих признаков соответствует распределению половых хромосом в процессе мейоза. Поскольку большинство генов, локализованных в Х-хромосоме, не имеют своих аллелей в Y-хромосоме, то у гетерогаметного пола (XY) в фенотипе проявляются все рецессивные гены, содержащиеся в их единственной Х-хромосоме. Гены, если они имеются в Y-хромосоме, проявляются также только у гетерогаметного пола.
Наследование признаков, определяемых генами, локализованными в Х и Y-хромосомах, называют сцепленным с полом. Впервые оно было описано Т. Морганом и его коллегами на примере рецессивного признака “white” — белые глаза.
Как видно из схемы, результаты прямого и обратного скрещиваний в случае наличия сцепления с полом разные. В прямом скрещивании гомозиготная красноглазая самка передает доминантный ген W и дочерям и сыновьям, благодаря чему все гибриды F 1 имеют красные глаза. Скрещивание гетерозиготных самок F 1 с самцами F 1 дает в F 2 только красноглазых самок, одна половина которых является гомозиготными, а другая — гетерозиготными. Среди самцов F 2 наблюдается расщепление на красноглазых и белоглазых в соотношении 1: 1, которое обусловлено гетерозиготностью самок F 1 , так как свою единственную Х-хромосому сыновья наследуют от матери. Общая формула расщепления по окраске глаз в F 2 (без учета пола) — 3: 1. На наличие сцепления признака с полом указывает то, что белая окраска глаз в F 2 проявляется только у самцов.

В обратном скрещивании рецессивная гомозиготная белоглазая самка передает ген w вместе с Х-хромосомой и дочерям и сыновьям F 1 , но проявляется он только у самцов. У самок F 1 этот ген подавляется доминантным аллельным геном, полученным от отца, и поэтому глаза у них красные. Таким образом, признак передается от отца к дочерям, а от матери к сыновьям. Такое наследование называется крисс-кросс (крест-накрест). Скрещивание самок и самцов F 1 дает мух двух фенотипических классов (красноглазых и белоглазых) в соотношении 1: 1, которое полностью соответствует распределению половых хромосом.
Описанный тип наследования окраски глаз у дрозофилы является закономерным для всех организмов в отношении признаков, которые определяются генами, локализованными в Х-хромосоме.

Сцепленное с полом наследование используется для ранней диагностики пола у животных, что важно для сельскохозяйственного производства. В птицеводстве важно определять пол “суточных” цыплят, чтобы ставить петушков и курочек на разный рацион, откармливая петушков на мясо. Для диагностики пола используется крисс-кросс наследование признака окраски пера. При скрещивании пестрой курицы (признак доминантный) с черным петухом (признак рецессивный) в F 1 все петушки, получившие доминантный ген от матери, будут пестрыми, а курочки — черными.
У человека сцепленно с полом наследуются такие наследственные аномалии, как гемофилия и дальтонизм. Поскольку у человека гетерогаметным является мужской пол, то эти аномалии проявляются, в основном, у мужчин. Женщины обычно являются носительницами таких генов, имея их в гетерозиготном состоянии.
При разведении тутового шелкопряда крисс-кросс наследование используется для отбора самцов по окраске грены (признак сцеплен с полом), так как выход шелка из коконов тутового шелкопряда мужского пола на 20-30% выше.
Картина сцепленного с полом наследования может искажаться, если наблюдаются отдельные случаи нерасхождения половых хромосом в процессе мейоза. Так, при скрещивании белоглазой самки дрозофилы с красноглазым самцом (см. выше схему наследования крисс-кросс) в F 1 , помимо красноглазых самок и белоглазых самцов, появляются единичные белоглазые самки и красноглазые самцы. Причиной этого отклонения является нерасхождение Х-хромосом у исходной самки. В процессе гаметогенеза в яйцеклетку попадает не одна Х-хромосома, а обе, или же, наоборот, ни одной, а обе попадают в полярное тельце. При оплодотворении таких яйцеклеток нормальными сперматозоидами и развиваются красноглазые самцы и белоглазые самки.
Потомство, которое образуется в результате первичного нерасхождения хромосом у самки, имеет разные, не соответствующие норме сочетания и количество половых хромосом. Однако, генетическая инертность Y-хромосомы делает особей с кариотипом ХХY женскими и жизнеспособными, а с кариотипом Х0 — мужскими и также жизнеспособными. Зиготы, не получившие Х-хромосомы (Y0 ), погибают, так же как (за редким исключением) и зиготы с тремя Х-хромосомами.
Схема наследования белой окраски глаз у дрозофилы (ген white)
при нерасхождении X-хромосом у самки

У дрозофилы выведена линия (double yellow — двойная желтая), у которой из поколения в поколение нарушается наследование сцепленного с полом признака — желтая окраска тела. У самок этой линии Х-хромосомы соединены друг с другом в проксимальной части и имеют одну центромеру. В связи с этим в мейозе они ведут себя как одна хромосома и в анафазе отходят к одному полюсу.
Гетерогаметность одного пола определяет соответствие соотношения полов в каждом поколении организмов формуле 1: 1. Это соотношение совпадает с расщеплением при анализирующем скрещивании. Рассмотрим его на примере дрозофилы, у которой определение пола соответствует Lygaeus-типу. Набор хромосом у дрозофилы состоит из трех пар аутосом и двух половых хромосом. Самка образует один тип гамет с гаплоидным набором (3A+X), а самец в равных количествах два типа гамет (3A+X) и (3A+Y). В итоге в следующем поколении развивается одинаковое количество самок и самцов.

Такое наследование наблюдается при разных типах хромосомного механизма определения пола, и вероятность рождения потомков мужского и женского пола в норме одинакова. Однако баланс полов может быть нарушен, если в половых хромосомах возникают летальные мутации. Рассмотрим случай, когда рецессивная летальная мутация (l ) возникла в одной из двух Х-хромосом самки дрозофилы (X Bl ), маркированной доминантной мутацией Bar (В ) — полосковидные глаза. Рассмотрите схему скрещивания такой самки с нормальным самцом дикого типа (+), имеющим круглые глаза.

Как видно из схемы, появление рецессивной летальной мутации в одной из Х-хромосом самки приводит к гибели половины мужского потомства. Об этом судят по отсутствию самцов с полосковидными глазами, получившими от матери Х-хромосому с летальным геном (X Bl ).
Гены, определяющие признаки пола, имеются не только в половых хромосомах, но и в аутосомах. С другой стороны, признаки, которые наследуются сцепленно с полом, часто не имеют прямого отношения к полу. Существует особая категория признаков, которые проявляются только у одного пола. Это — ограниченные полом признаки . Определяющие их гены имеются у обоих полов и могут находиться как в половых хромосомах, так и аутосомах. Однако работают эти гены, т.е. проявляют свое действие на уровне фенотипа, только у одного пола. К числу таких признаков относятся, например, молочность и жирность молока у коров, яйценоскость и размер яиц у кур. Эти признаки, которыми обладают особи женского пола, могут целиком определяться генотипом отца. Такое явление широко используется в селекции животных при использовании отцовских особей-производителей для получения высококачественного потомства.
Гены, определяющие развитие вторичных половых признаков, имеются как у мужчин, так и у женщин, но их проявление контролируется гормонами.
Пол может оказывать влияние на характер проявления признака, т.е. на его доминантность или рецессивность. В этом случае признаки называют зависимыми от пола . Так, например, у овец ген, определяющий развитие рогов, является доминантным у самцов и рецессивным — у самок. В связи с этим гетерозиготные самки являются комолыми, а гетерозиготные самцы — рогатыми. У человека точно так же наследуется признак плешивости. Зависимые от пола признаки находятся под сильным влиянием половых гормонов, соотношение которых может либо усилить, либо ослабить экспрессию гена.
Итак, подведем итог, касающийся механизма определения пола. Пол, как любой другой признак организма, детерминирован генетически. В определении пола у большинства животных и растений основная роль принадлежит половым хромосомам. Расщепление по полу соответствует соотношению 1: 1, что обусловлено равновероятным образованием двух типов гамет (1/2 с Х и 1/2 с Y хр.) у гетерогаметного пола (XY ). Гетерогаметным может быть как мужской, так и женский пол.
Определение пола — это начальный этап становления пола, за которым следует процесс его дифференциации, приводящий к развитию двух разных половых типов — женского и мужского. У животных половая дифференциация затрагивает всю организацию особи: строение органов размножения, внешнюю морфологию, обмен веществ, поведение, гормональный баланс, продолжительность жизни и пр. Половые различия которые обеспечивают комбинативную изменчивость внутри вида, а также его изоляцию, являются адаптивным механизмом.
Различают первичные и вторичные половые признаки. Первые непосредственно обеспечивают осуществление полового процесса. В частности, к ним относятся различия в строении внешних и внутренних половых органов женских и мужских особей. Развитие вторичных половых признаков является результатом нормального функционирования гонад (т.е. опосредовано первичными половыми признаками) и способствует половому размножению. Регулируется развитие вторичных половых признаков с помощью половых гормонов.
На процесс дифференциации пола оказывают влияние как генотипические факторы, так и внешняя среда.
Еще в начале ХХ в. было высказано предположение, что зигота является потенциально бисексуальной, но существуют механизмы, осуществляющие дифференциацию пола. Одним из таких механизмов является баланс половых хромосом и аутосом, при нарушении которого развитие пола отклоняется либо в сторону женского, либо в сторону мужского пола. Необходимость такого баланса впервые была установлена в опытах К. Бриджеса (лаборатория Т. Моргана), который обнаружил линию дрозофилы, дающую наряду с нормальными самцами и самками большой процент интерсексов. Интерсексы представляют собой смесь первичных и вторичных мужских и женских половых признаков, образуя все переходные типы: от сходных в основном с самцами до сходных с самками. Все они стерильны. В опыте Бриджеса они возникли в потомстве триплоидных самок, оплодотворенных нормальными диплоидными самцами, и содержали три набора аутосом и нормальное количество половых хромосом: 2Х+3А. Наряду с типичными интерсексами, в потомстве были представлены особи с гипертрофированными признаками женского пола — суперсамки (3Х+2А), и мужского пола — суперсамцы (XY+3X).
На основании этих результатов Бриджес пришел к выводу, что не само присутствие двух половых хромосом (XX или XY) определяет развитие пола, а баланс половых хромосом и гаплоидных наборов аутосом. Поскольку у дрозофилы Y-хромосома генетически инертна, то важно только количество Х-хромосом. Все особи с отношением 2Х: 2А = 1 являются самками, особи с отношением 1Х: 2А = 0,5 — самцами, типы с промежуточными между 1 и 0,5 отношениями являются интерсексами, а отношения больше 1 дают суперсамок, меньше 0,5 — суперсамцов.
Аномальное развитие пола при изменении числа наборов аутосом обусловлено нарушением баланса генов, которые участвуют в развитии пола. Поскольку гены проявляют свое действие в конкретных условиях, то на их функционирование оказывают влияние внешние факторы. Так, потомство триплоидных самок дрозофилы воспитывалось в условиях высокой и низкой температур. В обоих случаях развивались интерсексы, но при высокой температуре преимущественно с признаками самки, а при пониженной — с признаками самца. Таким образом, окончательное развитие пола является результатом сложных взаимодействий генов, локализованных как в половых хромосомах, так и в аутосомах, друг с другом и с факторами окружающей среды.
Изначальная бисексуальность зигот подтверждается фактами переопределения пола в процессе индивидуального развития. Классический пример — морской червь Bonellia viridis. Свободноплавающие личинки этого червя развиваются в самок. Если же личинка остается прикрепленной к материнской особи, из нее развивается самец. Будучи отделена от самки, такая личинка, начавшая развиваться в самца, изменяет направление дифференциации пола в женскую сторону и из нее развивается интерсекс. В хоботке самки имеются химические регуляторы, способные переопределять пол личинок.

Большой интерес представляет экспериментальное переопределение пола. Путем воздействия гормональными препаратами у ряда животных удается получить полное превращение пола вплоть до способности формировать половые клетки противоположного пола. Такое превращение известно у некоторых лягушек, рыб, птиц и других животных. Так, раннее удаление яичника у самок кур и голубей может изменить в мужскую сторону окраску оперения, поведение и даже вызвать развитие семенника. У крупного рогатого скота наблюдались случаи рождения разнополых двойнь, в которых бычок оказывался нормальным, а телка — стерильной, со многими чертами самцового типа. Такие двойни носят название “фримартинов”. Их появление обусловлено тем, что семенники мужского эмбриона рано начинают выделять мужской гормон, который попадает в кровь и оказывает влияние на близнеца.
Один из ярких примеров полного переопределения пола описан в 1953 г. японским ученым Т. Ямамото. Опыт проводился на белых и красных медаках (Oryzias latipes), у которых доминантный ген красной окраски находится в Y-хромосоме. При такой локализации гена при скрещивании самцы всегда будут красными, а самки — белыми. Фенотипических самцов кормили с добавлением в корм женского полового гормона. В результате оказалось, что все красные рыбки с генотипом самца являются самками с нормальными яичниками и женскими вторичными половыми признаками.
Переопределение пола может быть следствием мутаций отдельных генов, участвующих в дифференциации пола. Так, у дрозофилы в одной из аутосом обнаружен рецессивный ген tra , присутствие которого в гомозиготном состоянии обусловливает развитие женских зигот (XX) в фенотипических самцов, оказывающихся стерильными. Самцы XY, гомозиготные по этому гену, являются плодовитыми.
Аналогичные гены найдены у растений. Так, у кукурузы рецессивная мутация silkless в гомозиготном состоянии вызывает стерильность семяпочек, в связи с чем обоеполое растение функционирует как мужское. У сорго обнаружены два доминантных гена, комплементарное взаимодействие которых также вызывает женскую стерильность.
У наездника Habrobracon пол определяется по тому же типу, что и у пчел: диплоидные самки развиваются из оплодотворенных яиц, а гаплоидные самцы партеногенетически. Но иногда самцы могут развиваться из оплодотворенных яиц. Причина такой ситуации лежит в действии специфического гена, в гомозиготном состоянии определяющего развитие зиготы по мужскому типу.
Правильность хромосомной теории определения пола подтверждается существованием половых мозаиков, или гинандроморфов , совмещающих в себе части тела мужского и женского полов. Известны разные типы гинандроморфов: латеральные, переднезадние, мозаичные.

Билатеральный гинандроморф
Drosophila melanogaster
Латеральный гинандроморфизм описан у насекомых, у кур, у певчих птиц. В этом случае одна половина тела соответствует женскому типу, вторая — мужскому. При мозаичном гинандроморфизме большая часть тела имеет признаки одного пола, и лишь отдельные участки — признаки противоположного пола. Этот тип описан, в частности, у дрозофилы. Чаще всего причиной появления гинандроморфов является утрата одной из двух Х-хромосом в раннем дроблении зиготы с кариотипом самки (ХХ). Клетки с кариотипом Х0 обнаруживают признаки мужского пола. Чем раньше произойдет элиминация Х-хромосомы, тем больше участков мужского типа будет представлено в теле взрослой мухи. Обнаруживаются такие мозаики по проявлению рецессивных генов, которые в зиготе находились в гетерозиготном состоянии, но проявились фенотипически в клетках с кариотипом Х0.
Еще одной причиной гинандроморфизма может быть развитие зародыша из яйцеклетки с двумя ядрами (дизиготический гинандроморфизм). В этом случае мозаики могут быть соматическими, если оба ядра имеют один и тот же набор половых хромосом, но разный генотип (например, одно ядро Аа, а другое — аа), или половыми, если одно ядро ХХ, а другое ХY, или теми и другими одновременно. Подобный тип гинандроморфизма описан у шелковичного червя, бабочки, дрозофилы.
Известен также гинандроморфизм, причиной которого является полиспермия. Он обнаружен у дрозофилы. В яйцеклетке дрозофилы могут сформироваться два женских гаплоидных пронуклеуса, с одной Х-хромосомой каждый. При проникновении в яйцеклетку двух сперматозоидов один пронуклеус может оплодотвориться сперматозоидом с Х-хромосомой, а другой — сперматозоидом с Y-хромосомой. После первого дробления образуются два бластомера, один с кариотипом ХХ, другой — ХY, что в дальнейшем приведет к развитию гинандроморфа.
Хромосомы живых организмов делят на аутосомы и половые хромосомы . Последние также называют аллосомами , гоносомами .
В клетках большинства диплоидных организмов половых хромосом всего две. При этом у одного пола половые хромосомы одинаковы, т. е. гомологичны. У другого пола - половые хромосомы различаются между собой. Пол с одинаковыми аллосомами называется гомогаметным , пол с разными - гетерогаметным .
Гомогаметный пол образует одинаковые гаметы, гетерогаметный пол образует два типа гамет, различающиеся одной хромосомой. У одних групп организмов гетерогаметным является мужской пол (например, у млекопитающих), у других – гетерогаметен женский пол (например, у птиц).
Принято половые хромосомы обозначать латинскими буквами X, Y или Z, W. У женщин их набор XX, у мужчин - XY. У самок птиц аллосомы обозначают ZW, у самцов - ZZ.
В природе существуют виды, у которых гетерогаметный пол имеет лишь одну половую хромосому. В этом случае отсутствие второй обозначают цифрой 0, получается генотип X0. В этом случае гетерогаметный пол образует два типа гамет: одни содержат одну половую хромосому, другие - ни одной.
Следует отметить, что пол организмов не всегда определяется половыми хромосомами. Например, у крокодилов аллосом нет вообще, и пол развивающейся в яйце особи определяется температурой окружающей среды. Пол может зависеть от количества гаплоидных наборов. Так у некоторых насекомых (например, муравьев и пчел) из неоплодотворенных яиц (n) развиваются самцы, а из оплодотворенных (2n) - самки. При отсутствии половых хромосом пол определяется определенными аллелями генов, расположенных в аутосомах. Помимо перечисленных, в природе существуют другие, более редкие, механизмы детерминации (определения) пола.
Наследование, сцепленное с полом
В половых хромосомах находится немало генов, особенно в X-хромосоме. Многие из них определяют признаки, которые не связаны с детерминацией (развитием) пола. При этом эти признаки оказываются сцепленными с полом, и их наследование зависит от наследования пола. Наследование признаков, сцепленных с полом, совпадает с наследованием X- и Y-хромосом.
Наследование признаков, определяемые генами Y-хромосомы, всегда будет происходить только гетерогаметным полом (в случае млекопитающих - только самцами). В случае с X-хромосомой ситуация сложнее, так как данная хромосома присутствует в генотипах женских и мужских организмов. При этом самцы всегда получают ее от матери, а самки - от обоих родителей.
Наследование, сцепленное с полом, было замечено Т. Морганом и подтверждено проведенными им опытами на дрозофиле.
У дрозофил красные глаза (R) доминируют над белыми (r). Морган заметил, что при реципрокных скрещиваниях получаются разные результаты. (Реципрокные скрещивания: в одной части скрещиваний самка носитель признака, в другой части – самец.)
При скрещивании чистой линии красноглазых самок (RR) с белоглазыми самцами (по-идее rr) все потомство оказывалось красноглазым (по-идее Rr). Однако при скрещивании чистой линии красноглазых самцов (вроде как RR) с белоглазыми самками (rr) в потомстве все самцы оказывались белоглазыми (наследовали признак матери), а все самки - красноглазыми (наследовали признак отца). По-идее же все должны были быть красноглазыми по фенотипу (однако гетерозиготами Rr по генотипу).
Объяснение данному феномену можно было дать, предположив, что ген окраски глаз локализуется только в X-хромосоме. Тогда у самцов этот ген всегда находится в единичном экземпляре. Поскольку наследовать X-хромосому они могут только от самок-родительниц, то, если эта хромосома содержит рецессивный аллель, он неминуемо проявится, даже если их отцы были доминантными по этому признаку (самцы его просто не передают сыновьям). Но самцы передают его своим дочерям. Поэтому все самки-потомки от красноглазых самцов и белоглазых самок были красноглазыми.
Дальнейшие исследования Моргана по скрещиванию F 1 подтвердили, что признак наследуется как сцепленный с полом.
У человека с полом сцеплены такие заболевания как гемофилия и дальтонизм. Оба признака рецессивны и локализованы в X-хромосоме. У 50% сыновей матерей-гетерозигот эти заболевания проявятся. Если при этом болен и отец, то обладателями дефектного признака станет и 50% дочерей. Когда же болен отец, а мать - здоровая гомозигота, то все дети будут здоровы, однако все дочери будут носителями признака (так как окажутся гетерозиготами).